有丝分裂是真核生物细胞生命周期中的核心环节,对于遗传信息的传递和物种的繁殖具有至关重要的作用。在这一过程中,母细胞的遗传物质首先经过复制,随后被平均分配到两个子细胞中。这一精密的分配是通过纺锤体引导染色体定向、整列及分离而完成的。染色体的运动和分离是由位于着丝粒部位的动粒蛋白复合体与纺锤体微管之间的动态相互作用所调控的。尽管动粒与微管如何协同调控染色体运动是细胞生物学领域研究的热点问题,但在植物细胞中对这一机制的理解仍然相对有限。
近日,四川大学邓星光课题组在Nature Plants发表了题为A kinetochore-associated kinesin-7 motor cooperates with BUB3.3 to regulate mitotic chromosome congression in Arabidopsis thaliana的研究论文。该论文发现了一个新的微管马达蛋白调控植物细胞有丝分裂期染色体集会的机制。

该课题组之前在对拟南芥纺锤体组装检验点(SAC)的研究中发现BUB3.3蛋白的缺失突变体表现出独特的染色体排列异常现象,而其他SAC成员的突变体并未显示出这一细胞学特征(PNAS 2024, 121: e2322677121)。为了探究导致染色体排列缺陷的机制,该课题组对BUB3.3进行了体内纯化,并通过质谱分析鉴定到一个尚未报道过的微管驱动蛋白。初步研究显示,该驱动蛋白在细胞分裂过程中定位于动粒,因此将其命名为KAK1(Kinetochore-Associated Kinesin 1)。KAK1的缺失会导致部分染色体无法建立正确的双极定向,从而在有丝分裂中期不能整列到纺锤体中央。
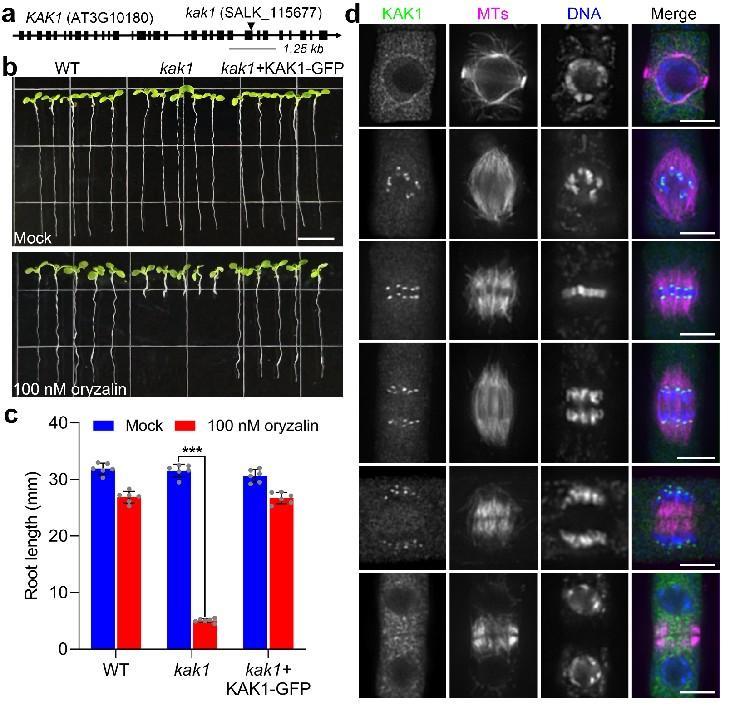
图1:KAK1是一个定位于动粒的驱动蛋白
尽管KAK1蛋白与BUB3.3蛋白之间存在相互作用,并且两者的缺失突变体都显示出有丝分裂中期染色体排列异常的表型,但进一步的研究揭示了它们在功能上的一些区别。具体来说,KAK1的缺失突变体(kak1)能够使有丝分裂中期暂停,直到所有的染色体都正确地排列在赤道板上。相比之下,BUB3.3的缺失突变体(bub3.3)则无法在有丝分裂中期实现这种暂停,导致其在后期经常出现染色体分离错误。此外,当在KAK1的缺失突变体中进一步突变SAC成员MAD1时,会导致其染色体在后期错误分离,这一现象与BUB3.3缺失突变体的表现相似。这些结果表明,KAK1主要调控有丝分裂过程中染色体的聚集,而并不直接参与SAC信号通路的调控。
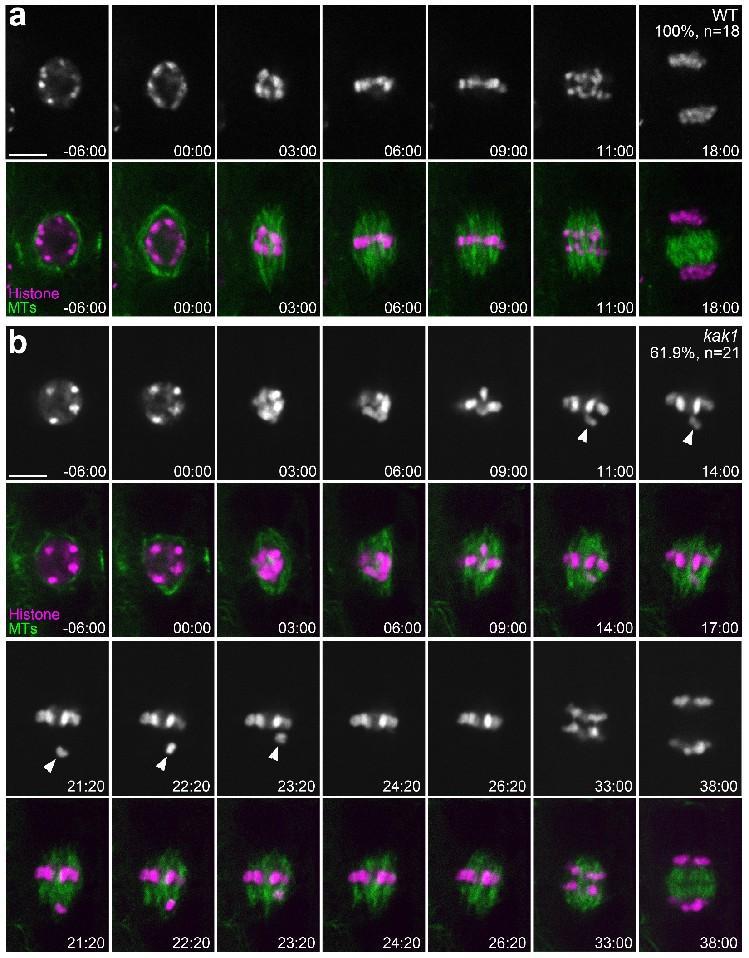
图2:KAK1缺失导致染色体整列异常
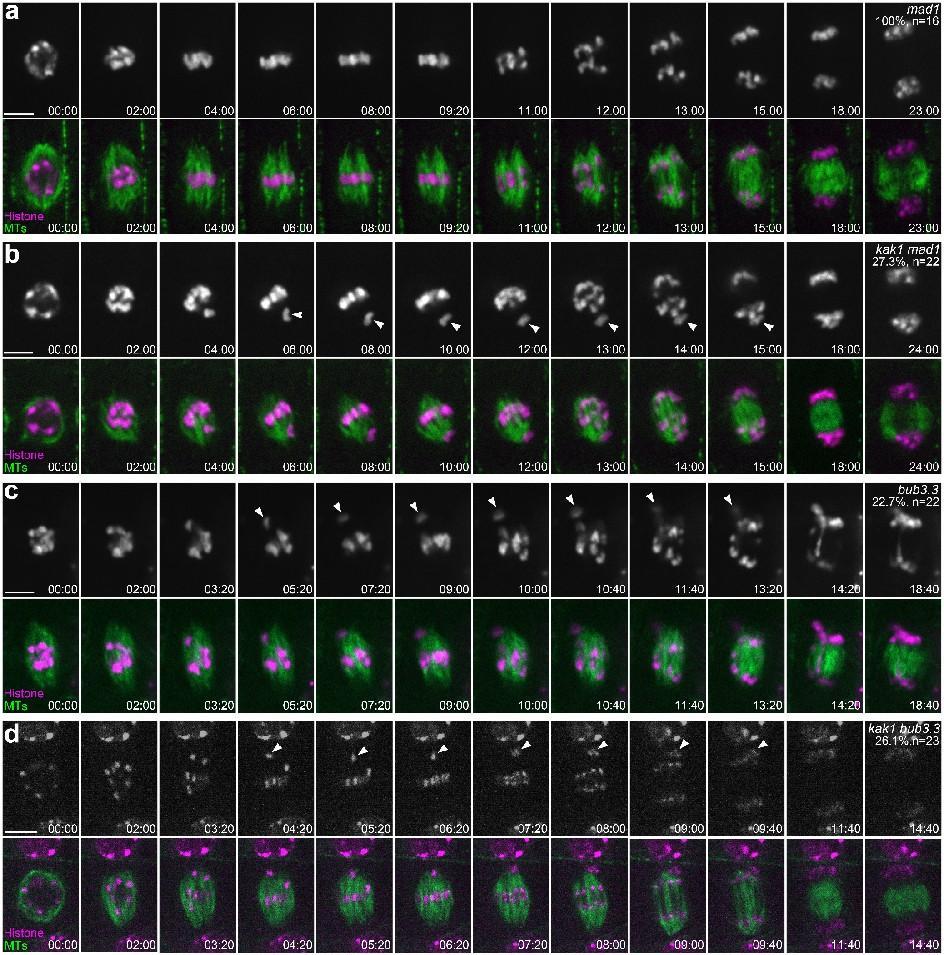
图3:抑制SAC导致kak1发生染色体错误分离
进一步研究发现,KAK1与BUB3.3之间存在相互依赖的关系,它们共同协作以确保动粒的定位。将KAK1人工锚定在动粒上可以恢复bub3.3突变体染色体整列缺陷的表型。反之将BUB3.3人工锚定到动粒上却不能恢复kak1突变体的表型。这表明BUB3.3作用于KAK1的上游,以影响KAK1在动粒上定位的方式参与染色体集会的调控。此外,点突变和截短实验分析发现KAK1的马达区域虽然对其执行功能是必需的,但并不影响其在动粒上的定位。
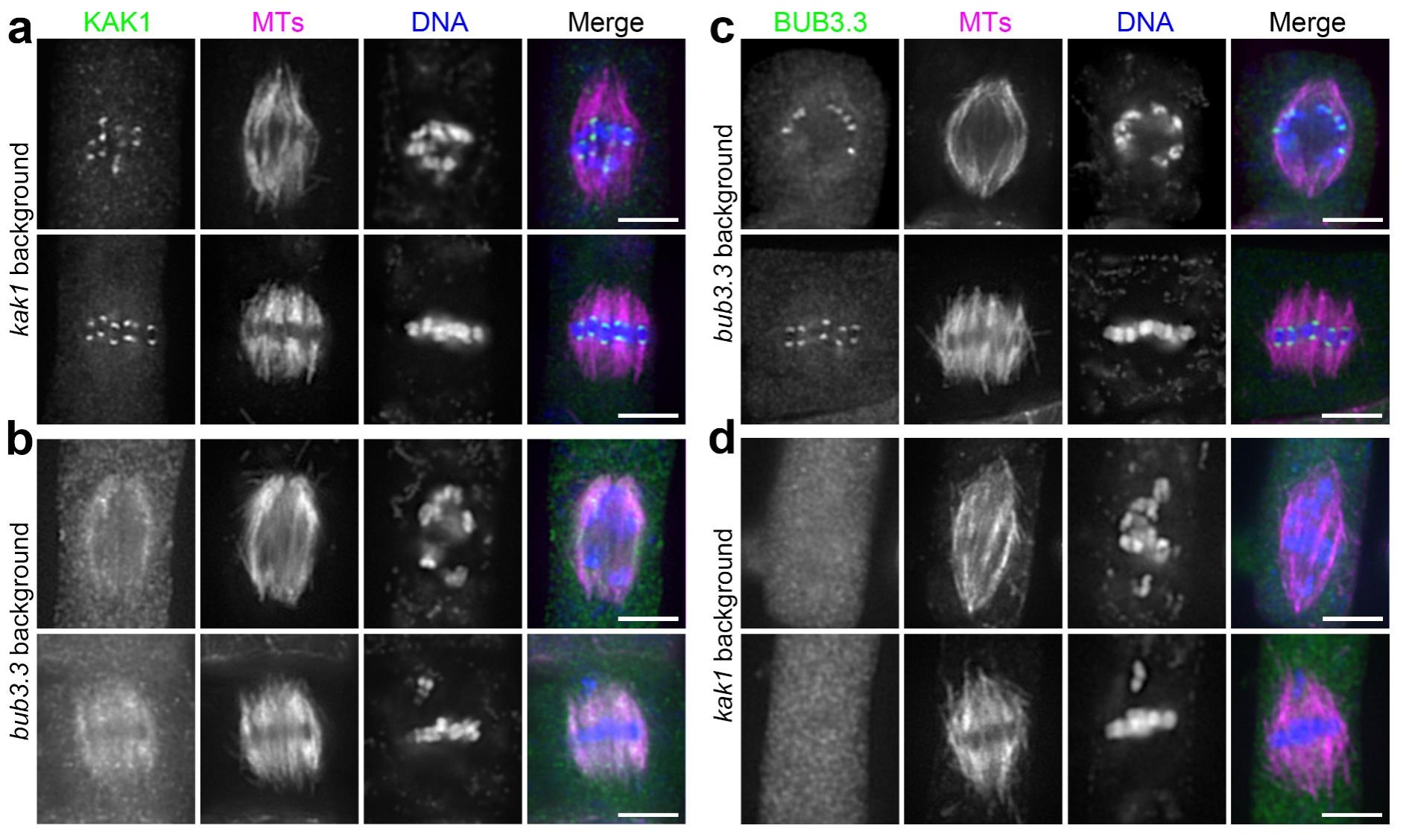
图4:KAK1与BUB3.3相互依赖完成动粒定位
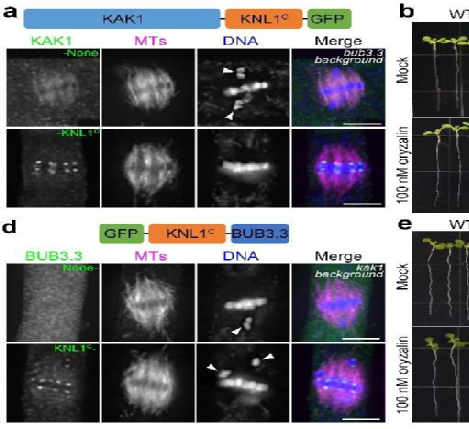
图5:KAK1在染色体集会过程中作用于BUB3.3下游
综上所述,该研究发现了一个定位在动粒上的植物驱动蛋白KAK1,揭示了其在SAC蛋白BUB3.3调控下参与染色体的集会过程。这项工作不仅增进了我们对植物细胞有丝分裂调控机制的理解,而且为植物细胞工程和遗传改良提供了重要的科学基础。
四川大学在读博士研究生唐小雅为该论文第一作者,邓星光副教授为通讯作者。四川大学林宏辉教授和加州大学柳波教授为该研究提供了重要的支持和指导。该研究得到了国家自然科学基金以及四川省自然科学基金等项目的资助。
论文链接:https://www.nature.com/articles/s41477-024-01824-7


